Definition von Antikörpern – Isotope, Struktur, Funktionen, Medizinische Anwendungen und vieles mehr
Frage
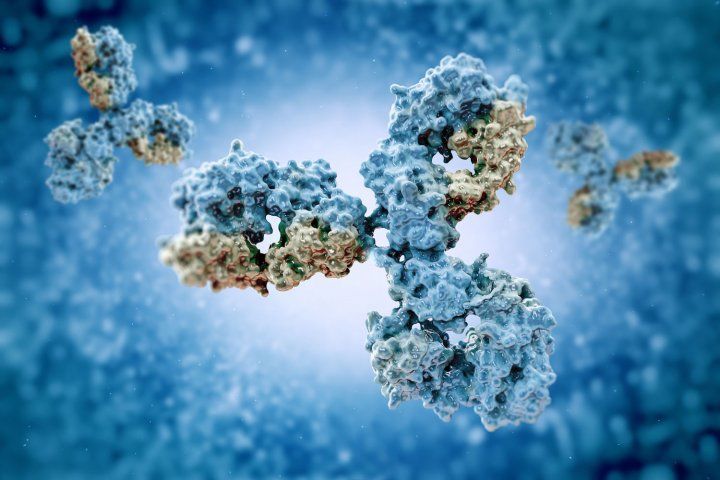
Antikörper sind das Rückgrat des Immunsystems im menschlichen Körper,In diesem Artikel würden wir uns eingehend mit der Definition von Antikörpern befassen,ihre Isotope,medizinische Anwendung von Antikörpern und vieles mehr.
Ein Antikörper (Ab), auch bekannt als Immunoglobulin (Ich G),es ist groß, Y-förmiges Protein, das hauptsächlich von Plasmazellen produziert wird und vom Immunsystem zur Neutralisierung von Krankheitserregern wie pathogenen Bakterien und Viren verwendet wird.
Der Antikörper erkennt ein einzigartiges Molekül des Erregers, ein Antigen genannt, über das Fragment Antigen-Bindung (Fab) variable Region wie SARS-CoV-2, das Virus, das COVID-19 verursacht. Sie bekämpfen Infektionen, indem sie Teile des Virus blockieren, die zur Infektion einer Zelle benötigt werden, oder indem sie sie für die Zerstörung durch das Immunsystem markieren.
Antikörper werden von Immunzellen produziert, die als B-Zellen bekannt sind. Die unglaubliche Auswahl an Antikörpern, die wir produzieren können, beruht auf der unglaublichen Auswahl an B-Zellen, die wir haben. Wenn wir mit einem Virus infiziert sind, Ein kleiner Satz von B-Zellen erkennt das Virus und, über ein paar Wochen, mit Hilfe anderer Immunzellen, die als T-Zellen bekannt sind, Sie lernen, immer stärkere Antikörper gegen das Virus zu produzieren. Diese B-Zellen reifen und vermehren sich zu Fabriken für die Antikörperproduktion, die als Plasmazellen bekannt sind.
Jede Spitze des “Y.” eines Antikörpers enthält ein Paratop (analog zu einem Schloss) das ist spezifisch für ein bestimmtes Epitop (analog zu einem Schlüssel) auf einem Antigen, Dadurch können diese beiden Strukturen präzise miteinander verbunden werden.
Y-förmige Antikörper
Verwendung dieses Bindungsmechanismus, ein Antikörper kann Etikett eine Mikrobe oder eine infizierte Zelle zum Angriff durch andere Teile des Immunsystems, oder kann sein Ziel direkt neutralisieren (zum Beispiel, durch Hemmung eines Teils einer Mikrobe, der für ihre Invasion und ihr Überleben wesentlich ist).
Abhängig vom Antigen, Die Bindung kann den biologischen Prozess, der die Krankheit verursacht, behindern oder Makrophagen aktivieren, um die Fremdsubstanz zu zerstören.
Die Fähigkeit eines Antikörpers, mit den anderen Komponenten des Immunsystems zu kommunizieren, wird über seine Fc-Region vermittelt (befindet sich am Fuße des “Y.”), Diese enthält eine konservierte Glykosylierungsstelle, die an diesen Wechselwirkungen beteiligt ist. Die Produktion von Antikörpern ist die Hauptfunktion des humoralen Immunsystems.
Antikörper sind Glykoproteine, die zur Immunglobulin-Superfamilie gehören. Sie machen den größten Teil der Gammaglobulinfraktion der Blutproteine aus. Sie bestehen typischerweise aus grundlegenden Struktureinheiten - jede mit zwei großen schweren Ketten und zwei kleinen leichten Ketten.
Es gibt verschiedene Arten von schweren Antikörperketten, die die fünf verschiedenen Arten von kristallisierbaren Fragmenten definieren (Fc) das kann an die Antigen-bindenden Fragmente gebunden sein.
Die fünf verschiedenen Arten von Fc-Regionen ermöglichen die Gruppierung von Antikörpern in fünf Isotypen. Jede Fc-Region eines bestimmten Antikörper-Isotyps kann an seinen spezifischen Fc-Rezeptor binden (FcR), außer IgD, Das ist im Wesentlichen die BCR, Auf diese Weise kann der Antigen-Antikörper-Komplex je nach gebundenem FcR unterschiedliche Rollen vermitteln.
Die Fähigkeit eines Antikörpers, an sein entsprechendes FcR zu binden, wird durch die Struktur des Glykans weiter moduliert(s) an konservierten Standorten innerhalb seiner Fc-Region vorhanden.
Die Fähigkeit von Antikörpern, an FcRs zu binden, hilft dabei, die geeignete Immunantwort für jede Art von Fremdkörper zu steuern, auf die sie treffen. Zum Beispiel, IgE ist verantwortlich für eine allergische Reaktion bestehend aus Mastzelldegranulation und Histaminfreisetzung.
Das Fab-Paratop von IgE bindet an allergisches Antigen, zum Beispiel Hausstaubmilbenpartikel, während seine Fc-Region an den Fc-Rezeptor ε bindet. Die Allergen-IgE-FcRε-Wechselwirkung vermittelt die allergische Signalübertragung, um Erkrankungen wie Asthma zu induzieren.
Obwohl die allgemeine Struktur aller Antikörper sehr ähnlich ist, Eine kleine Region an der Spitze des Proteins ist extrem variabel, Millionen von Antikörpern mit leicht unterschiedlichen Spitzenstrukturen, oder Antigen-Bindungsstellen, existieren. Diese Region ist bekannt als die hypervariable Region.
Jede dieser Varianten kann an ein anderes Antigen binden. Diese enorme Vielfalt an Antikörperparatopen auf den Antigen-bindenden Fragmenten ermöglicht es dem Immunsystem, eine ebenso große Vielfalt an Antigenen zu erkennen.
Die große und vielfältige Population von Antikörperparatopen wird durch zufällige Rekombinationsereignisse einer Reihe von Gensegmenten erzeugt, die für verschiedene Antigen-Bindungsstellen kodieren (oder Paratope), gefolgt von zufälligen Mutationen in diesem Bereich des Antikörpergens, die weitere Vielfalt schaffen.
Dieser Rekombinationsprozess, der klonale Antikörper-Paratop-Diversität erzeugt, wird als V bezeichnet(D)J- oder VJ-Rekombination. Das Antikörperparatop ist polygen, bestehend aus drei Genen, V., D, und J.. Jeder Paratopenort ist auch polymorph, so dass während der Antikörperproduktion, ein Allel von V., einer von D., und einer von J wird gewählt.
Diese Gensegmente werden dann unter Verwendung einer zufälligen genetischen Rekombination zusammengefügt, um das Paratop herzustellen. Die Regionen, in denen die Gene zufällig zusammen rekombiniert werden, sind die hypervariablen Regionen, die verwendet werden, um verschiedene Antigene auf klonaler Basis zu erkennen.
Antikörpergene organisieren sich auch in einem Prozess neu, der als Klassenwechsel bezeichnet wird und den einen Typ des Fc-Fragments der schweren Kette in einen anderen ändert, Erzeugen eines anderen Isotyps des Antikörpers, der die Antigen-spezifische variable Region beibehält. Dies ermöglicht die Verwendung eines einzelnen Antikörpers durch verschiedene Arten von Fc-Rezeptoren, auf verschiedenen Teilen des Immunsystems ausgedrückt.
Isotope von Antikörpern
Die membrangebundene Form eines Antikörpers kann als a bezeichnet werden Oberflächen-Immunglobulin (SIG) oder ein Membran-Immunglobulin (mIg).
Es ist Teil der B-Zell-Rezeptor (BCR), Dadurch kann eine B-Zelle erkennen, wann ein bestimmtes Antigen im Körper vorhanden ist, und die Aktivierung der B-Zellen auslösen
.Die BCR besteht aus oberflächengebundenen IgD- oder IgM-Antikörpern und assoziierten Ig- & agr; - und Ig- & bgr; -Heterodimeren, die zur Signalübertragung fähig sind. Eine typische menschliche B-Zelle wird haben 50,000 zu 100,000 an seine Oberfläche gebundene Antikörper.
Bei Antigenbindung, Sie gruppieren sich in großen Patches, was überschreiten kann 1 Mikrometer im Durchmesser, auf Lipidflößen, die die BCRs von den meisten anderen Zellsignalrezeptoren isolieren.
Diese Pflaster können die Effizienz der zellulären Immunantwort verbessern. Beim Menschen, Die Zelloberfläche ist mehrere hundert Nanometer lang um die B-Zell-Rezeptoren herum freigelegt,Dies isoliert die BCRs weiter von konkurrierenden Einflüssen.
Antikörper oder Immunglobuline gibt es in verschiedenen Formen. Basierend auf Unterschieden in den Aminosäuresequenzen im konstanten Bereich der schweren Ketten werden sie weiter in fünf Klassen eingeteilt. Diese sind:
- IgG – mit gamma schwerer Kette
- IgM - enthält mu schwere Kette
- IgA – Alpha schwere Kette enthaltend
- IgD-haltige schwere Delta-Kette
- IgE-haltige Epsilon-Schwerkette
Sie sind jeweils mit einem benannt “Ich G” Präfix, das für Immunglobulin steht (Ein Name, der manchmal austauschbar mit Antikörpern verwendet wird) und unterscheiden sich in ihren biologischen Eigenschaften, funktionelle Orte und Fähigkeit, mit verschiedenen Antigenen umzugehen, wie in der Tabelle dargestellt.
Die verschiedenen Suffixe der Antikörper-Isotypen bezeichnen die verschiedenen Arten von schweren Ketten, die der Antikörper enthält, mit jeder Klasse der schweren Kette alphabetisch benannt: a (Alpha), c (Gamma), d (Delta), e (Epsilon), und μ (mu). Dies führt zu IgA, IgG, IgD, IgE, und IgM, beziehungsweise.
Struktur
Antikörper sind schwer (~ 150 kDa) globuläre Plasmaproteine. Die Größe eines Antikörpermoleküls beträgt etwa 10 nm.Sie haben Zuckerketten (Glykane) zu konservierten Aminosäureresten gegeben.
Mit anderen Worten, Antikörper sind Glykoproteine.Die gebundenen Glykane sind für die Struktur und Funktion des Antikörpers von entscheidender Bedeutung. Unter anderem können die exprimierten Glykane die Affinität eines Antikörpers zu seinem entsprechenden FcR modulieren(s).

Struktur eines Antikörpers
Die grundlegende funktionelle Einheit jedes Antikörpers ist ein Immunglobulin (Ich G) Monomer (enthält nur eine Ig-Einheit); sekretierte Antikörper können ebenso wie IgA mit zwei Ig-Einheiten dimer sein, tetramer mit vier Ig-Einheiten wie Knochenfische IgM, oder pentamer mit fünf Ig-Einheiten, wie Säugetier IgM.
Mehrere Immunglobulindomänen bilden die beiden schweren Ketten (rot und Blau) und die zwei leichten Ketten (grün und gelb) eines Antikörpers. Die Immunglobulindomänen bestehen aus zwischen 7 (für konstante Domänen) und 9 (für variable Domänen) β-Stränge.
Die variablen Teile eines Antikörpers sind seine V-Regionen, und der konstante Teil ist seine C-Region.
Immunglobulindomänen
Das Ig-Monomer ist a “Y.”-geformtes Molekül, das aus vier Polypeptidketten besteht; zwei identisch schwere Ketten und zwei identische leichte Ketten verbunden durch Disulfidbrücken.
Jede Kette besteht aus strukturellen Domänen, die als Immunglobulindomänen bezeichnet werden. Diese Domänen enthalten etwa 70–110 Aminosäuren und werden in verschiedene Kategorien eingeteilt (zum Beispiel, variabel oder IgV, und konstant oder IgC) entsprechend ihrer Größe und Funktion.
Sie haben eine charakteristische Immunglobulinfalte, in der zwei Beta-Blätter eine bilden “Sandwich” gestalten, durch Wechselwirkungen zwischen konservierten Cysteinen und anderen geladenen Aminosäuren zusammengehalten.
Schwere Kette
Es gibt fünf Arten von schwerer Ig-Kette von Säugetieren, die mit den griechischen Buchstaben bezeichnet werden: a, d, e, c, und μ. Die Art der vorhandenen schweren Kette definiert die Klasse von Antikörper; Diese Ketten sind in IgA zu finden, IgD, IgE, IgG, und IgM-Antikörper, beziehungsweise.
Deutliche schwere Ketten unterscheiden sich in Größe und Zusammensetzung; α und γ enthalten ungefähr 450 Aminosäuren, während μ und ε ungefähr 550 Aminosäuren.
Jede schwere Kette hat zwei Regionen, das konstante Region und das variable Region. Die konstante Region ist bei allen Antikörpern des gleichen Isotyps identisch, unterscheidet sich jedoch in Antikörpern verschiedener Isotypen.
Schwere Ketten γ, α und δ haben einen konstanten Bereich bestehend aus drei Tandem (in einer Linie) Ig-Domänen, und einen Scharnierbereich für zusätzliche Flexibilität;schwere Ketten μ und ε haben einen konstanten Bereich aus vier Immunglobulindomänen.
Die variable Region der schweren Kette unterscheidet sich in Antikörpern, die von verschiedenen B-Zellen produziert werden, Dies ist jedoch für alle Antikörper gleich, die von einer einzelnen B-Zelle oder einem B-Zell-Klon produziert werden. Der variable Bereich jeder schweren Kette beträgt ungefähr 110 Aminosäuren lang und besteht aus einer einzelnen Ig-Domäne.
Lichterkette
Bei Säugetieren gibt es zwei Arten der leichten Kette von Immunglobulin, die Lambda genannt werden (λ) und Kappa (Herr).Eine leichte Kette hat zwei aufeinanderfolgende Domänen: eine konstante Domäne und eine variable Domäne.
Die ungefähre Länge einer leichten Kette beträgt 211 zu 217 Aminosäuren. Jeder Antikörper enthält zwei leichte Ketten, die immer identisch sind; nur eine Art von leichter Kette, κ oder λ, ist pro Antikörper in Säugetieren vorhanden. Andere Arten von leichten Ketten, wie das iota (j) Kette, kommen in anderen Wirbeltieren wie Haien vor (Chondrichthyes) und knöcherne Fische (Teleostei).
CDRs, Fv, Fab- und Fc-Regionen
Verschiedene Teile eines Antikörpers haben unterschiedliche Funktionen. Speziell, das “Waffen” (die sind in der Regel identisch) Stellen enthalten, die an bestimmte Moleküle binden können, Ermöglichen der Erkennung spezifischer Antigene.
Diese Region des Antikörpers wird als bezeichnet Fab (Fragment, Antigen-Bindung) Region. Es besteht aus einer konstanten und einer variablen Domäne aus jeder schweren und leichten Kette des Antikörpers.
Das Paratop am aminoterminalen Ende des Antikörpermonomers wird durch die variablen Domänen der schweren und leichten Ketten geformt. Die variable Domäne wird auch als F bezeichnetV. Region und ist die wichtigste Region für die Bindung an Antigene.
Um genau zu sein, variable Schleifen von β-Strängen, je drei am Licht (V.L) und schwer (V.H) Ketten sind für die Bindung an das Antigen verantwortlich.
Diese Schleifen werden als komplementaritätsbestimmende Regionen bezeichnet (CDRs). Die Strukturen dieser CDRs wurden von Chothia et al. Und in jüngerer Zeit von North et al. Und Nikoloudis et al.
Im Rahmen der Immunnetzwerktheorie, CDRs werden auch als Idiotypen bezeichnet. Nach der Theorie des Immunnetzwerks, Das adaptive Immunsystem wird durch Wechselwirkungen zwischen Idiotypen reguliert.
Die Basis des Y spielt eine Rolle bei der Modulation der Immunzellaktivität. Diese Region heißt die Fc (Fragment, kristallisierbar) Region, und besteht aus zwei schweren Ketten, die je nach Klasse des Antikörpers zwei oder drei konstante Domänen beisteuern.
Somit, Die Fc-Region stellt sicher, dass jeder Antikörper eine geeignete Immunantwort für ein bestimmtes Antigen erzeugt, durch Bindung an eine bestimmte Klasse von Fc-Rezeptoren, und andere Immunmoleküle, wie Komplementproteine.
Auf diese Weise, es vermittelt verschiedene physiologische Wirkungen, einschließlich der Erkennung opsonisierter Partikel (Bindung an FcγR), Lyse von Zellen (Bindung an Komplement), und Degranulation von Mastzellen, Basophile, und Eosinophile (Bindung an FcεR).
Zusammenfassend, Die Fab-Region des Antikörpers bestimmt die Antigenspezifität, während die Fc-Region des Antikörpers den Klasseneffekt des Antikörpers bestimmt.
Da nur die konstanten Domänen der schweren Ketten die Fc-Region eines Antikörpers bilden, Die Klassen der schweren Kette in Antikörpern bestimmen ihre Klasseneffekte. Mögliche Klassen schwerer Ketten in Antikörpern umfassen Alpha, Gamma, Delta, Epsilon, und mu, und sie definieren die Isotypen des Antikörpers IgA, G, D, E, und M, beziehungsweise.
Dies impliziert, dass unterschiedliche Isotypen von Antikörpern aufgrund ihrer unterschiedlichen Fc-Regionen, die unterschiedliche Rezeptortypen binden und aktivieren, unterschiedliche Klasseneffekte haben.
Mögliche Klasseneffekte von Antikörpern umfassen: Opsonisierung, Agglutination, Hämolyse, Komplementaktivierung, Degranulation der Mastzellen, und Neutralisierung (obwohl dieser Klasseneffekt eher durch die Fab-Region als durch die Fc-Region vermittelt werden kann).
Dies impliziert auch, dass Fab-vermittelte Effekte auf Mikroben oder Toxine gerichtet sind, während Fc-vermittelte Effekte auf Effektorzellen oder Effektormoleküle gerichtet sind.
Funktionen
Die Hauptkategorien der Antikörperwirkung umfassen die folgenden:
- Neutralisation, bei denen neutralisierende Antikörper Teile der Oberfläche einer Bakterienzelle oder eines Virions blockieren, um dessen Angriff unwirksam zu machen
- Agglutination, in welchen Antikörpern “zusammenkleben” Fremdzellen zu Klumpen, die attraktive Ziele für die Phagozytose sind
- Niederschlag, in welchen Antikörpern “zusammenkleben” Serumlösliche Antigene, Sie werden gezwungen, in Klumpen, die attraktive Ziele für die Phagozytose sind, aus der Lösung auszufallen
- Komplementaktivierung (Fixierung), bei denen Antikörper, die an eine fremde Zelle gebunden sind, das Komplement dazu anregen, es mit einem Membranangriffskomplex anzugreifen, was zu folgendem führt:
- Lyse der Fremdzelle
- Förderung der Entzündung durch chemotaktische Anziehung von Entzündungszellen
Aktivierte B-Zellen differenzieren entweder in Antikörper produzierende Zellen, sogenannte Plasmazellen, die lösliche Antikörper sezernieren, oder in Gedächtniszellen, die jahrelang im Körper überleben, damit sich das Immunsystem an ein Antigen erinnern und bei zukünftigen Expositionen schneller reagieren kann.
In der pränatalen und neonatalen Lebensphase, Das Vorhandensein von Antikörpern wird durch passive Immunisierung der Mutter sichergestellt. Die frühe endogene Antikörperproduktion variiert für verschiedene Arten von Antikörpern, und erscheinen in der Regel innerhalb der ersten Lebensjahre.
Da Antikörper frei im Blut vorhanden sind, Sie sollen Teil des humoralen Immunsystems sein. Zirkulierende Antikörper werden von klonalen B-Zellen produziert, die spezifisch nur auf ein Antigen reagieren (Ein Beispiel ist ein Virus-Kapsid-Protein-Fragment).
Antikörper tragen auf drei Arten zur Immunität bei: Sie verhindern, dass Krankheitserreger in Zellen eindringen oder diese schädigen, indem sie an sie binden; Sie stimulieren die Entfernung von Krankheitserregern durch Makrophagen und andere Zellen, indem sie den Krankheitserreger beschichten; und sie lösen die Zerstörung von Krankheitserregern aus, indem sie andere Immunantworten wie den Komplementweg stimulieren.
Antikörper lösen auch eine vasoaktive Amindegranulation aus, um zur Immunität gegen bestimmte Arten von Antigenen beizutragen (Helminthen, Allergene).
Aktivierung des Komplements
Antikörper, die an Oberflächenantigene binden (zum Beispiel, auf Bakterien) ziehen die erste Komponente der Komplementkaskade mit ihrer Fc-Region an und initiieren die Aktivierung der “klassisch” Komplementsystem.
Dies führt auf zwei Arten zur Abtötung von Bakterien, Die Bindung der Antikörper- und Komplementmoleküle markiert die Mikrobe für die Aufnahme durch Phagozyten in einem als Opsonisierung bezeichneten Prozess; Diese Phagozyten werden von bestimmten Komplementmolekülen angezogen, die in der Komplementkaskade erzeugt werden.
Zweite, Einige Komponenten des Komplementsystems bilden einen Membranangriffskomplex, der Antikörpern hilft, das Bakterium direkt abzutöten (Bakteriolyse).
Aktivierung von Effektorzellen
Zur Bekämpfung von Krankheitserregern, die sich außerhalb der Zellen vermehren, Antikörper binden an Krankheitserreger, um sie miteinander zu verbinden, was dazu führt, dass sie agglutinieren.
Da ein Antikörper mindestens zwei Paratope hat, Es kann mehr als ein Antigen binden, indem es identische Epitope bindet, die auf den Oberflächen dieser Antigene getragen werden.
Durch Beschichten des Erregers, Antikörper stimulieren Effektorfunktionen gegen den Erreger in Zellen, die ihre Fc-Region erkennen.
Diejenigen Zellen, die beschichtete Pathogene erkennen, haben Fc-Rezeptoren, welche, wie der Name schon sagt, interagieren mit der Fc-Region von IgA, IgG, und IgE-Antikörper.
Die Bindung eines bestimmten Antikörpers an den Fc-Rezeptor an einer bestimmten Zelle löst eine Effektorfunktion dieser Zelle aus; Phagozyten werden phagozytieren, Mastzellen und Neutrophile degranulieren, Natürliche Killerzellen setzen Zytokine und zytotoxische Moleküle frei; Dies führt letztendlich zur Zerstörung der eindringenden Mikrobe.
Die Aktivierung natürlicher Killerzellen durch Antikörper initiiert einen zytotoxischen Mechanismus, der als antikörperabhängige zellvermittelte Zytotoxizität bekannt ist (ADCC) - Dieser Prozess könnte die Wirksamkeit von monoklonalen Antikörpern erklären, die in biologischen Therapien gegen Krebs eingesetzt werden.
Die Fc-Rezeptoren sind isotypspezifisch, Dies gibt dem Immunsystem mehr Flexibilität, nur die geeigneten Immunmechanismen für verschiedene Krankheitserreger aufrufen.
Natürliche Antikörper
Menschen und höhere Primaten produzieren ebenfalls “natürliche Antikörper” die vor einer Virusinfektion im Serum vorhanden sind. Natürliche Antikörper wurden als Antikörper definiert, die ohne vorherige Infektion hergestellt werden, Impfung, andere Fremdantigenexposition oder passive Immunisierung.
Diese Antikörper können den klassischen Komplementweg aktivieren, der zur Lyse umhüllter Viruspartikel führt, lange bevor die adaptive Immunantwort aktiviert wird.
Viele natürliche Antikörper sind gegen das Disaccharid Galactose α gerichtet(1,3)-Galaktose (α-Gal), welches als terminaler Zucker auf glykosylierten Zelloberflächenproteinen gefunden wird, und als Reaktion auf die Produktion dieses Zuckers durch im menschlichen Darm enthaltene Bakterien erzeugt.
Es wird angenommen, dass xenotransplantierte Organe abgestoßen werden, teils, das Ergebnis von natürlichen Antikörpern, die im Serum des Empfängers zirkulieren und an auf dem Spendergewebe exprimierte α-Gal-Antigene binden
Immunglobulin-Diversität
Praktisch alle Mikroben können eine Antikörperantwort auslösen. Die erfolgreiche Erkennung und Ausrottung vieler verschiedener Arten von Mikroben erfordert die Vielfalt der Antikörper; Ihre Aminosäurezusammensetzung variiert, so dass sie mit vielen verschiedenen Antigenen interagieren können.
Es wurde geschätzt, dass der Mensch etwa 10 Milliarden verschiedene Antikörper, jedes kann ein bestimmtes Epitop eines Antigens binden.
Obwohl ein riesiges Repertoire an verschiedenen Antikörpern in einem einzelnen Individuum erzeugt wird, Die Anzahl der zur Herstellung dieser Proteine verfügbaren Gene ist durch die Größe des menschlichen Genoms begrenzt.
Es haben sich mehrere komplexe genetische Mechanismen entwickelt, die es Wirbeltier-B-Zellen ermöglichen, aus einer relativ kleinen Anzahl von Antikörpergenen einen vielfältigen Pool von Antikörpern zu erzeugen.
Domänenvariabilität
Die chromosomale Region, die einen Antikörper codiert, ist groß und enthält mehrere unterschiedliche Genorte für jede Domäne des Antikörpers - die Chromosomenregion, die Gene der schweren Kette enthält (IGH @) wird auf Chromosom gefunden 14, und die Loci, die Gene der leichten Kette von Lambda und Kappa enthalten (IGL @ und IGK @) finden sich auf Chromosomen 22 und 2 in Menschen.
Eine dieser Domänen wird als variable Domäne bezeichnet, welches in jeder schweren und leichten Kette jedes Antikörpers vorhanden ist, kann sich aber in verschiedenen Antikörpern unterscheiden, die aus verschiedenen B-Zellen erzeugt werden.
Unterschiede, zwischen den variablen Domänen, befinden sich in drei Schleifen, die als hypervariable Regionen bekannt sind (HV-1, HV-2 und HV-3) oder komplementaritätsbestimmende Regionen (CDR1, CDR2 und CDR3). CDRs werden innerhalb der variablen Domänen von konservierten Framework-Regionen unterstützt.
Der Ort der schweren Kette enthält ungefähr 65 verschiedene Gene mit variabler Domäne, die sich alle in ihren CDRs unterscheiden. Die Kombination dieser Gene mit einer Reihe von Genen für andere Domänen des Antikörpers erzeugt eine große Kavallerie von Antikörpern mit einem hohen Grad an Variabilität.
Diese Kombination heißt V.(D)J Rekombination diskutiert unten.
V.(D)J Rekombination
Somatische Rekombination von Immunglobulinen, auch bekannt als V.(D)J Rekombination, beinhaltet die Erzeugung einer einzigartigen variablen Immunglobulinregion.
Die variable Region jeder schweren oder leichten Immunglobulinkette ist in mehreren Teilen kodiert - sogenannte Gensegmente (Subgene). Diese Segmente werden als variabel bezeichnet (V.), Vielfalt (D) und beitreten (J.) Segmente.
V., D- und J-Segmente finden sich in schweren Ig-Ketten, In leichten Ig-Ketten finden sich jedoch nur V- und J-Segmente. Mehrere Kopien des V., D- und J-Gensegmente existieren, und sind im Genom von Säugetieren tandemartig angeordnet. Im Knochenmark, Jede sich entwickelnde B-Zelle wird eine variable Region des Immunglobulins durch zufällige Auswahl und Kombination eines V zusammensetzen, ein D- und ein J-Gensegment (oder ein V- und ein J-Segment in der leichten Kette).
Da es mehrere Kopien von jedem Typ von Gensegment gibt, und verschiedene Kombinationen von Gensegmenten können verwendet werden, um jede variable Region des Immunglobulins zu erzeugen, Dieser Prozess erzeugt eine große Anzahl von Antikörpern, jeweils mit unterschiedlichen Paratopen, und damit unterschiedliche Antigenspezifitäten.
Die Umlagerung mehrerer Subgene (d.h.. V2 Familie) für Lambda leichte Kette ist Immunglobulin mit der Aktivierung von microRNA miR-650 gekoppelt, was die Biologie von B-Zellen weiter beeinflusst.
RAG-Proteine spielen bei V eine wichtige Rolle(D)J Rekombination beim Schneiden von DNA in einer bestimmten Region. Ohne das Vorhandensein dieser Proteine, V.(D)Eine J-Rekombination würde nicht auftreten.
Nachdem eine B-Zelle während V ein funktionelles Immunglobulin-Gen produziert(D)J Rekombination, Es kann keine andere variable Region ausdrücken (Ein Prozess, der als allelischer Ausschluss bekannt ist) Somit kann jede B-Zelle Antikörper produzieren, die nur eine Art variabler Kette enthalten.
Somatische Hypermutation und Affinitätsreifung
Nach Aktivierung mit Antigen, B-Zellen beginnen sich schnell zu vermehren. In diesen sich schnell teilenden Zellen, Die Gene, die die variablen Domänen der schweren und leichten Ketten codieren, unterliegen einer hohen Rate an Punktmutationen, durch einen Prozess namens somatische Hypermutation (SHM).
SHM führt zu ungefähr einer Nukleotidänderung pro variablem Gen., pro Zellteilung. Als Folge, Alle Tochter-B-Zellen erwerben leichte Aminosäureunterschiede in den variablen Domänen ihrer Antikörperketten.
Dies dient dazu, die Diversität des Antikörperpools zu erhöhen und die Antigen-Bindungsaffinität des Antikörpers zu beeinflussen.
Einige Punktmutationen führen zur Produktion von Antikörpern mit einer schwächeren Wechselwirkung (geringe Affinität) mit ihrem Antigen als der ursprüngliche Antikörper, und einige Mutationen erzeugen Antikörper mit einer stärkeren Wechselwirkung (hohe Affinität).
B-Zellen, die Antikörper mit hoher Affinität auf ihrer Oberfläche exprimieren, erhalten während Wechselwirkungen mit anderen Zellen ein starkes Überlebenssignal, während diejenigen mit Antikörpern mit niedriger Affinität dies nicht tun, und wird durch Apoptose sterben.
Somit, B-Zellen, die Antikörper mit einer höheren Affinität für das Antigen exprimieren, werden diejenigen mit schwächeren Affinitäten für Funktion und Überleben übertreffen, wodurch die durchschnittliche Affinität von Antikörpern mit der Zeit zunehmen kann.
Der Prozess der Erzeugung von Antikörpern mit erhöhten Bindungsaffinitäten wird genannt Affinitätsreifung. Die Affinitätsreifung erfolgt in reifen B-Zellen nach V.(D)J Rekombination, und ist auf die Hilfe von Helfer-T-Zellen angewiesen.
Klassenwechsel
Isotyp- oder Klassenwechsel ist ein biologischer Prozess, der nach Aktivierung der B-Zelle stattfindet, Dies ermöglicht es der Zelle, verschiedene Klassen von Antikörpern zu produzieren (IgA, IgE, oder IgG).
Die verschiedenen Klassen von Antikörpern, und damit Effektorfunktionen, werden durch die Konstante definiert (C) Regionen der schweren Kette des Immunglobulins.
Anfänglich, naive B-Zellen exprimieren nur Zelloberflächen-IgM und IgD mit identischen Antigen-Bindungsregionen. Jeder Isotyp ist für eine bestimmte Funktion angepasst; deshalb, nach der Aktivierung, ein Antikörper mit einem IgG, IgA, oder eine IgE-Effektorfunktion könnte erforderlich sein, um ein Antigen effektiv zu eliminieren.
Durch Klassenwechsel können verschiedene Tochterzellen derselben aktivierten B-Zelle Antikörper mit unterschiedlichen Isotypen produzieren.
Während des Klassenwechsels ändert sich nur die konstante Region der schweren Kette des Antikörpers; die variablen Regionen, und daher Antigenspezifität, bleiben unverändert.
Somit kann die Nachkommenschaft einer einzelnen B-Zelle Antikörper produzieren, alle spezifisch für das gleiche Antigen, aber mit der Fähigkeit, die Effektorfunktion zu erzeugen, die für jede antigene Herausforderung geeignet ist.
Der Klassenwechsel wird durch Zytokine ausgelöst; Der erzeugte Isotyp hängt davon ab, welche Zytokine in der B-Zell-Umgebung vorhanden sind.
Der Klassenwechsel erfolgt im Genort der schweren Kette durch einen Mechanismus, der als Klassenwechselrekombination bezeichnet wird (CSR). Dieser Mechanismus beruht auf konservierten Nukleotidmotiven, namens Schalter (S) Regionen, gefunden in der DNA stromaufwärts von jedem Gen der konstanten Region (außer in der δ-Kette).
Der DNA-Strang wird durch die Aktivität einer Reihe von Enzymen an zwei ausgewählten S-Regionen gebrochen.
Das Exon der variablen Domäne wird durch einen Prozess, der als nicht homologe Endverbindung bezeichnet wird, wieder verbunden (NHEJ) auf den gewünschten konstanten Bereich (c, α oder ε). Dieser Prozess führt zu einem Immunglobulin-Gen, das einen Antikörper eines anderen Isotyps codiert.
Spezifitätsbezeichnungen
Ein Antikörper kann aufgerufen werden monospezifisch wenn es eine Spezifität für dasselbe Antigen oder Epitop aufweist,oder bispezifisch, wenn sie eine Affinität zu zwei verschiedenen Antigenen oder zwei verschiedenen Epitopen auf demselben Antigen aufweisen.
Eine Gruppe von Antikörpern kann aufgerufen werden polyvalent (oder unspezifisch) wenn sie eine Affinität zu verschiedenen Antigenen oder Mikroorganismen haben. Intravenöses Immunglobulin, wenn nicht anders angegeben, besteht aus einer Vielzahl verschiedener IgG (polyklonales IgG). Im Gegensatz, monoklonale Antikörper sind identische Antikörper, die von einer einzelnen B-Zelle produziert werden.
Asymmetrische Antikörper
Heterodimere Antikörper, die auch asymmetrisch und Antikörper sind, ermöglichen eine größere Flexibilität und neue Formate für die Anbringung einer Vielzahl von Arzneimitteln an den Antikörperarmen.
Eines der allgemeinen Formate für einen heterodimeren Antikörper ist das “Knöpfe in Löcher” Format. Dieses Format ist spezifisch für den Teil der schweren Kette der konstanten Region in Antikörpern.
Das “Knöpfe” Teil wird hergestellt, indem eine kleine Aminosäure durch eine größere ersetzt wird. Es passt in die “Loch”, Dies wird durch Ersetzen einer großen Aminosäure durch eine kleinere entwickelt.
Was verbindet die “Knöpfe” zum “Löcher” sind die Disulfidbindungen zwischen jeder Kette. Das “Knöpfe in Löcher” Form erleichtert die antikörperabhängige zellvermittelte Zytotoxizität.
Einzelkettenvariable Fragmente (scFv) sind über ein kurzes Linkerpeptid mit der variablen Domäne der schweren und leichten Kette verbunden. Der Linker ist reich an Glycin, das gibt es mehr Flexibilität, und Serin / Threonin, das gibt es Spezifität.
Zwei verschiedene scFv-Fragmente können miteinander verbunden werden, über einen Scharnierbereich, auf die konstante Domäne der schweren Kette oder die konstante Domäne der leichten Kette. Dies gibt dem Antikörper Bispezifität, unter Berücksichtigung der Bindungsspezifitäten von zwei verschiedenen Antigenen.
Das “Knöpfe in Löcher” Das Format verbessert die Heterodimerbildung, unterdrückt jedoch nicht die Homodimerbildung.
Weitere Verbesserung der Funktion heterodimerer Antikörper, Viele Wissenschaftler suchen nach künstlichen Konstrukten.
Künstliche Antikörper sind weitgehend unterschiedliche Proteinmotive, die die funktionelle Strategie des Antikörpermoleküls nutzen, Sie sind jedoch nicht durch die Schleifen- und Gerüststrukturbeschränkungen des natürlichen Antikörpers begrenzt.
Die Möglichkeit, das kombinatorische Design der Sequenz und des dreidimensionalen Raums zu steuern, könnte das natürliche Design überschreiten und die Anbringung verschiedener Wirkstoffkombinationen an den Armen ermöglichen.
Heterodimere Antikörper haben eine größere Bandbreite an Formen, die sie annehmen können, und die an den Armen angebrachten Medikamente müssen nicht an jedem Arm gleich sein, Ermöglichen der Verwendung verschiedener Arzneimittelkombinationen bei der Krebsbehandlung.
Pharmazeutika sind in der Lage, hochfunktionelle bispezifische Produkte herzustellen, und sogar multispezifisch, Antikörper. Das Ausmaß, in dem sie funktionieren können, ist beeindruckend, da eine solche Änderung der Form von der natürlichen Form zu einer verminderten Funktionalität führen sollte.
Medizinische Anwendungen
Krankheitsdiagnose
Der Nachweis bestimmter Antikörper ist eine sehr häufige Form der medizinischen Diagnostik, und Anwendungen wie die Serologie hängen von diesen Methoden ab.
Beispielsweise, in biochemischen Tests zur Diagnose von Krankheiten,Aus dem Blut wird ein Titer von Antikörpern geschätzt, die gegen das Epstein-Barr-Virus oder die Lyme-Borreliose gerichtet sind.
Wenn diese Antikörper nicht vorhanden sind, entweder ist die Person nicht infiziert oder die Infektion ist aufgetreten a sehr vor langer Zeit, und die B-Zellen, die diese spezifischen Antikörper erzeugen, sind auf natürliche Weise zerfallen.

medizinische Diagnose von Antikörpern
In der klinischen Immunologie, Die Spiegel der einzelnen Klassen von Immunglobulinen werden durch Nephelometrie gemessen (oder Trübungsmessung) zur Charakterisierung des Antikörperprofils des Patienten. Erhöhungen in verschiedenen Klassen von Immunglobulinen sind manchmal nützlich, um die Ursache von Leberschäden bei Patienten zu bestimmen, für die die Diagnose unklar ist.[1] Beispielsweise, Erhöhtes IgA weist auf eine alkoholische Zirrhose hin, Erhöhtes IgM weist auf Virushepatitis und primäre biliäre Zirrhose hin, während IgG bei viraler Hepatitis erhöht ist, Autoimmunhepatitis und Leberzirrhose.
Autoimmunerkrankungen lassen sich häufig auf Antikörper zurückführen, die körpereigene Epitope binden; Viele können durch Blutuntersuchungen nachgewiesen werden. Antikörper, die gegen Antigene der Oberfläche roter Blutkörperchen bei immunvermittelter hämolytischer Anämie gerichtet sind, werden mit dem Coombs-Test nachgewiesen. Der Coombs-Test wird auch für das Antikörperscreening bei der Bluttransfusionspräparation und auch für das Antikörperscreening bei vorgeburtlichen Frauen verwendet.
Praktisch, Zur Diagnose von Infektionskrankheiten werden verschiedene immundiagnostische Methoden verwendet, die auf dem Nachweis komplexer Antigen-Antikörper beruhen, zum Beispiel ELISA, Immunfluoreszenz, Westlicher Fleck, Immundiffusion, Immunelektrophorese, und magnetischer Immunoassay.
Antikörper gegen humanes Choriongonadotropin werden in rezeptfreien Schwangerschaftstests verwendet.
Neue Dioxaborolan-Chemie ermöglicht radioaktives Fluorid (18F) Markierung von Antikörpern, Dies ermöglicht die Positronenemissionstomographie (HAUSTIER) Bildgebung von Krebs.
Krankheitstherapie
Eine gezielte monoklonale Antikörpertherapie wird zur Behandlung von Krankheiten wie rheumatoider Arthritis eingesetzt,Multiple Sklerose,Schuppenflechte,und viele Formen von Krebs, einschließlich Non-Hodgkin-Lymphom,Darmkrebs, Kopf-Hals-Krebs und Brustkrebs.
Einige Immunschwächen, wie X-chromosomale Agammaglobulinämie und Hypogammaglobulinämie, führen zu einem teilweisen oder vollständigen Mangel an Antikörpern. Diese Krankheiten werden häufig durch Induzieren einer kurzfristigen Form der Immunität behandelt, die als passive Immunität bezeichnet wird. Passive Immunität wird durch den Transfer von vorgefertigten Antikörpern in Form von menschlichem oder tierischem Serum erreicht, gepooltes Immunglobulin oder monoklonale Antikörper, in die betroffene Person.
Vorgeburtliche Therapie
Rhesusfaktor, auch als Rh D-Antigen bekannt, ist ein Antigen, das auf roten Blutkörperchen gefunden wird; Personen, die Rh-positiv sind (Rh +) haben dieses Antigen auf ihren roten Blutkörperchen und Individuen, die Rh-negativ sind (Rh–) unterlassen Sie.
Während der normalen Geburt, Geburtstrauma oder Komplikationen während der Schwangerschaft, Blut von einem Fötus kann in das System der Mutter gelangen.
Im Falle einer Rh-inkompatiblen Mutter und eines Kindes, Eine nachfolgende Blutmischung kann einen Rh sensibilisieren- Mutter des Rh-Antigens auf den Blutzellen des Rh + -Kindes, den Rest der Schwangerschaft setzen, und alle nachfolgenden Schwangerschaften, gefährdet für hämolytische Erkrankungen des Neugeborenen.
Rho(D) Immunglobulin-Antikörper sind spezifisch für menschliches RhD-Antigen. Anti-RhD-Antikörper werden als Teil eines vorgeburtlichen Behandlungsschemas verabreicht, um eine Sensibilisierung zu verhindern, die auftreten kann, wenn eine Rh-negative Mutter einen Rh-positiven Fötus hat.
Die Behandlung einer Mutter mit Anti-RhD-Antikörpern vor und unmittelbar nach Trauma und Entbindung zerstört das Rh-Antigen im System der Mutter vom Fötus.
Es ist wichtig zu beachten, dass dies geschieht, bevor das Antigen mütterliche B-Zellen dazu stimulieren kann “merken” Rh-Antigen durch Erzeugung von Gedächtnis-B-Zellen.
Deshalb, Ihr humorales Immunsystem wird keine Anti-Rh-Antikörper produzieren, und greift die Rh-Antigene des aktuellen oder nachfolgenden Babys nicht an.
Rho(D) Die Behandlung mit Immunglobulin verhindert eine Sensibilisierung, die zur Rh-Krankheit führen kann, verhindert oder behandelt jedoch nicht die Grunderkrankung selbst.
Kredit:
https://en.wikipedia.org/wiki/Antibody#Forms
Lassen Sie eine Antwort
Sie müssen Anmeldung oder registrieren um eine neue Antwort hinzuzufügen.