Antibodies Definitiion – Isotopes, Structure, Functions, Medical Applications And Lots More
Question
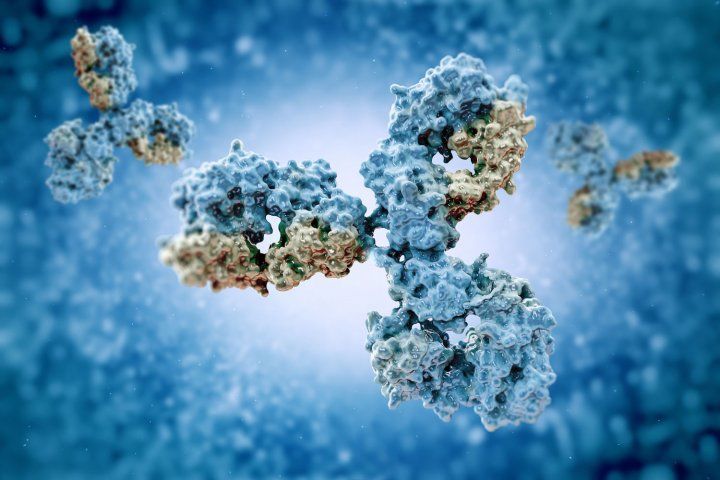
Antibodies are the backbone of the immune system in the human body,in this article we would take an in-depth look at antibodies definition,their isotopes,medical application of antibodies and lots more.
An antibody (Ab), also known as an immunoglobulin (Ig),is a large, Y-shaped protein produced mainly by plasma cells that is used by the immune system to neutralize pathogens such as pathogenic bacteria and viruses.
The antibody recognizes a unique molecule of the pathogen, called an antigen, via the fragment antigen-binding (Fab) variable region such as SARS-CoV-2, the virus that causes COVID-19. They fight infection by blocking parts of the virus needed to infect a cell or by marking them for destruction by the immune system.
Antibodies are produced by immune cells known as B cells. The incredible range of antibodies we can produce stem from the incredible range of B cells we have. When we are infected with a virus, a small set of B cells recognizes the virus and, over a couple of weeks, with the help of other immune cells known as T cells, they learn to produce stronger and stronger antibodies to the virus. These B cells mature and multiply into factories for antibody production known as plasma cells.
Each tip of the “Y” of an antibody contains a paratope (analogous to a lock) that is specific for one particular epitope (analogous to a key) on an antigen, allowing these two structures to bind together with precision.
Y-shaped antibodies
Using this binding mechanism, an antibody can tag a microbe or an infected cell for attack by other parts of the immune system, or can neutralize its target directly (for example, by inhibiting a part of a microbe that is essential for its invasion and survival).
Depending on the antigen, the binding may impede the biological process causing the disease or may activate macrophages to destroy the foreign substance.
The ability of an antibody to communicate with the other components of the immune system is mediated via its Fc region (located at the base of the “Y”), which contains a conserved glycosylation site involved in these interactions.The production of antibodies is the main function of the humoral immune system.
Antibodies are glycoproteins belonging to the immunoglobulin superfamily.They constitute most of the gamma globulin fraction of the blood proteins. They are typically made of basic structural units—each with two large heavy chains and two small light chains.
There are several different types of antibody heavy chains that define the five different types of crystallisable fragments (Fc) that may be attached to the antigen-binding fragments.
The five different types of Fc regions allow antibodies to be grouped into five isotypes. Each Fc region of a particular antibody isotype is able to bind to its specific Fc Receptor (FcR), except for IgD, which is essentially the BCR, thus allowing the antigen-antibody complex to mediate different roles depending on which FcR it binds.
The ability of an antibody to bind to its corresponding FcR is further modulated by the structure of the glycan(s) present at conserved sites within its Fc region.
The ability of antibodies to bind to FcRs helps to direct the appropriate immune response for each different type of foreign object they encounter.For example, IgE is responsible for an allergic response consisting of mast cell degranulation and histamine release.
IgE’s Fab paratope binds to allergic antigen, for example house dust mite particles, while its Fc region binds to Fc receptor ε. The allergen-IgE-FcRε interaction mediates allergic signal transduction to induce conditions such as asthma.
Though the general structure of all antibodies is very similar, a small region at the tip of the protein is extremely variable, allowing millions of antibodies with slightly different tip structures, or antigen-binding sites, to exist. This region is known as the hypervariable region.
Each of these variants can bind to a different antigen.This enormous diversity of antibody paratopes on the antigen-binding fragments allows the immune system to recognize an equally wide variety of antigens.
The large and diverse population of antibody paratope is generated by random recombination events of a set of gene segments that encode different antigen-binding sites (or paratopes), followed by random mutations in this area of the antibody gene, which create further diversity.
This recombinational process that produces clonal antibody paratope diversity is called V(D)J or VJ recombination. The antibody paratope is polygenic, made up of three genes, V, D, and J. Each paratope locus is also polymorphic, such that during antibody production, one allele of V, one of D, and one of J is chosen.
These gene segments are then joined together using random genetic recombination to produce the paratope. The regions where the genes are randomly recombined together is the hypervariable region used to recognise different antigens on a clonal basis.
Antibody genes also re-organize in a process called class switching that changes the one type of heavy chain Fc fragment to another, creating a different isotype of the antibody that retains the antigen-specific variable region. This allows a single antibody to be used by different types of Fc receptors, expressed on different parts of the immune system.
Isotopes Of Antibodies
The membrane-bound form of an antibody may be called a surface immunoglobulin (sIg) or a membrane immunoglobulin (mIg).
It is part of the B cell receptor (BCR), which allows a B cell to detect when a specific antigen is present in the body and triggers B cell activation
.The BCR is composed of surface-bound IgD or IgM antibodies and associated Ig-α and Ig-β heterodimers, which are capable of signal transduction.A typical human B cell will have 50,000 to 100,000 antibodies bound to its surface.
Upon antigen binding, they cluster in large patches, which can exceed 1 micrometer in diameter, on lipid rafts that isolate the BCRs from most other cell signaling receptors.
These patches may improve the efficiency of the cellular immune response.In humans, the cell surface is bare around the B cell receptors for several hundred nanometers,which further isolates the BCRs from competing influences.
Antibodies or immunoglobulins come in a variety of forms. Based on differences in the amino acid sequences at the constant region of the heavy chains they are further classified into five classes. These are:
- IgG – containing gamma heavy chain
- IgM – containing mu heavy chain
- IgA – containing alpha heavy chain
- IgD – containing delta heavy chain
- IgE – containing epsilon heavy chain
They are each named with an “Ig” prefix that stands for immunoglobulin (a name sometimes used interchangeably with antibody) and differ in their biological properties, functional locations and ability to deal with different antigens, as depicted in the table.
The different suffixes of the antibody isotypes denote the different types of heavy chains the antibody contains, with each heavy chain class named alphabetically: α (alpha), γ (gamma), δ (delta), ε (epsilon), and μ (mu). This gives rise to IgA, IgG, IgD, IgE, and IgM, respectively.
Structure
Antibodies are heavy (~150 kDa) globular plasma proteins. The size of an antibody molecule is about 10 nm.They have sugar chains (glycans) added to conserved amino acid residues.
In other words, antibodies are glycoproteins.The attached glycans are critically important to the structure and function of the antibody.Among other things the expressed glycans can modulate an antibody’s affinity for its corresponding FcR(s).

structure of an antibody
The basic functional unit of each antibody is an immunoglobulin (Ig) monomer (containing only one Ig unit); secreted antibodies can also be dimeric with two Ig units as with IgA, tetrameric with four Ig units like teleost fish IgM, or pentameric with five Ig units, like mammalian IgM.
Several immunoglobulin domains make up the two heavy chains (red and blue) and the two light chains (green and yellow) of an antibody. The immunoglobulin domains are composed of between 7 (for constant domains) and 9 (for variable domains) β-strands.
The variable parts of an antibody are its V regions, and the constant part is its C region.
Immunoglobulin domains
The Ig monomer is a “Y”-shaped molecule that consists of four polypeptide chains; two identical heavy chains and two identical light chains connected by disulfide bonds.
Each chain is composed of structural domains called immunoglobulin domains. These domains contain about 70–110 amino acids and are classified into different categories (for example, variable or IgV, and constant or IgC) according to their size and function.
They have a characteristic immunoglobulin fold in which two beta sheets create a “sandwich” shape, held together by interactions between conserved cysteines and other charged amino acids.
Heavy chain
There are five types of mammalian Ig heavy chain denoted by the Greek letters: α, δ, ε, γ, and μ.The type of heavy chain present defines the class of antibody; these chains are found in IgA, IgD, IgE, IgG, and IgM antibodies, respectively.
Distinct heavy chains differ in size and composition; α and γ contain approximately 450 amino acids, whereas μ and ε have approximately 550 amino acids.
Each heavy chain has two regions, the constant region and the variable region. The constant region is identical in all antibodies of the same isotype, but differs in antibodies of different isotypes.
Heavy chains γ, α and δ have a constant region composed of three tandem (in a line) Ig domains, and a hinge region for added flexibility;heavy chains μ and ε have a constant region composed of four immunoglobulin domains.
The variable region of the heavy chain differs in antibodies produced by different B cells, but is the same for all antibodies produced by a single B cell or B cell clone. The variable region of each heavy chain is approximately 110 amino acids long and is composed of a single Ig domain.
Light chain
In mammals there are two types of immunoglobulin light chain, which are called lambda (λ) and kappa (κ).A light chain has two successive domains: one constant domain and one variable domain.
The approximate length of a light chain is 211 to 217 amino acids.Each antibody contains two light chains that are always identical; only one type of light chain, κ or λ, is present per antibody in mammals. Other types of light chains, such as the iota (ι) chain, are found in other vertebrates like sharks (Chondrichthyes) and bony fishes (Teleostei).
CDRs, Fv, Fab and Fc regions
Different parts of an antibody have different functions. Specifically, the “arms” (which are generally identical) contain sites that can bind to specific molecules, enabling recognition of specific antigens.
This region of the antibody is called the Fab (fragment, antigen-binding) region. It is composed of one constant and one variable domain from each heavy and light chain of the antibody.
The paratope at the amino terminal end of the antibody monomer is shaped by the variable domains from the heavy and light chains. The variable domain is also referred to as the FV region and is the most important region for binding to antigens.
To be specific, variable loops of β-strands, three each on the light (VL) and heavy (VH) chains are responsible for binding to the antigen.
These loops are referred to as the complementarity-determining regions (CDRs). The structures of these CDRs have been clustered and classified by Chothia et al and more recently by North et al and Nikoloudis et al.
In the framework of the immune network theory, CDRs are also called idiotypes. According to immune network theory, the adaptive immune system is regulated by interactions between idiotypes.
The base of the Y plays a role in modulating immune cell activity. This region is called the Fc (Fragment, crystallizable) region, and is composed of two heavy chains that contribute two or three constant domains depending on the class of the antibody.
Thus, the Fc region ensures that each antibody generates an appropriate immune response for a given antigen, by binding to a specific class of Fc receptors, and other immune molecules, such as complement proteins.
By doing this, it mediates different physiological effects, including recognition of opsonized particles (binding to FcγR), lysis of cells (binding to complement), and degranulation of mast cells, basophils, and eosinophils (binding to FcεR).
In summary, the Fab region of the antibody determines antigen specificity while the Fc region of the antibody determines the antibody’s class effect.
Since only the constant domains of the heavy chains make up the Fc region of an antibody, the classes of heavy chain in antibodies determine their class effects. Possible classes of heavy chains in antibodies include alpha, gamma, delta, epsilon, and mu, and they define the antibody’s isotypes IgA, G, D, E, and M, respectively.
This implies different isotypes of antibodies have different class effects due to their different Fc regions binding and activating different types of receptors.
Possible class effects of antibodies include: Opsonisation, agglutination, haemolysis, complement activation, mast cell degranulation, and neutralisation (though this class effect may be mediated by the Fab region rather than the Fc region).
It also implies that Fab-mediated effects are directed at microbes or toxins, whilst Fc mediated effects are directed at effector cells or effector molecules.
Functions
The main categories of antibody action include the following:
- Neutralisation, in which neutralizing antibodies block parts of the surface of a bacterial cell or virion to render its attack ineffective
- Agglutination, in which antibodies “glue together” foreign cells into clumps that are attractive targets for phagocytosis
- Precipitation, in which antibodies “glue together” serum-soluble antigens, forcing them to precipitate out of solution in clumps that are attractive targets for phagocytosis
- Complement activation (fixation), in which antibodies that are latched onto a foreign cell encourage complement to attack it with a membrane attack complex, which leads to the following:
- Lysis of the foreign cell
- Encouragement of inflammation by chemotactically attracting inflammatory cells
Activated B cells differentiate into either antibody-producing cells called plasma cells that secrete soluble antibody or memory cells that survive in the body for years afterward in order to allow the immune system to remember an antigen and respond faster upon future exposures.
At the prenatal and neonatal stages of life, the presence of antibodies is provided by passive immunization from the mother. Early endogenous antibody production varies for different kinds of antibodies, and usually appear within the first years of life.
Since antibodies exist freely in the bloodstream, they are said to be part of the humoral immune system. Circulating antibodies are produced by clonal B cells that specifically respond to only one antigen (an example is a virus capsid protein fragment).
Antibodies contribute to immunity in three ways: They prevent pathogens from entering or damaging cells by binding to them; they stimulate removal of pathogens by macrophages and other cells by coating the pathogen; and they trigger destruction of pathogens by stimulating other immune responses such as the complement pathway.
Antibodies will also trigger vasoactive amine degranulation to contribute to immunity against certain types of antigens (helminths, allergens).
Activation of complement
Antibodies that bind to surface antigens (for example, on bacteria) will attract the first component of the complement cascade with their Fc region and initiate activation of the “classical” complement system.
This results in the killing of bacteria in two ways.First, the binding of the antibody and complement molecules marks the microbe for ingestion by phagocytes in a process called opsonization; these phagocytes are attracted by certain complement molecules generated in the complement cascade.
Second, some complement system components form a membrane attack complex to assist antibodies to kill the bacterium directly (bacteriolysis).
Activation of effector cells
To combat pathogens that replicate outside cells, antibodies bind to pathogens to link them together, causing them to agglutinate.
Since an antibody has at least two paratopes, it can bind more than one antigen by binding identical epitopes carried on the surfaces of these antigens.
By coating the pathogen, antibodies stimulate effector functions against the pathogen in cells that recognize their Fc region.
Those cells that recognize coated pathogens have Fc receptors, which, as the name suggests, interact with the Fc region of IgA, IgG, and IgE antibodies.
The engagement of a particular antibody with the Fc receptor on a particular cell triggers an effector function of that cell; phagocytes will phagocytose, mast cells and neutrophils will degranulate, natural killer cells will release cytokines and cytotoxic molecules; that will ultimately result in destruction of the invading microbe.
The activation of natural killer cells by antibodies initiates a cytotoxic mechanism known as antibody-dependent cell-mediated cytotoxicity (ADCC) – this process may explain the efficacy of monoclonal antibodies used in biological therapies against cancer.
The Fc receptors are isotype-specific, which gives greater flexibility to the immune system, invoking only the appropriate immune mechanisms for distinct pathogens.
Natural antibodies
Humans and higher primates also produce “natural antibodies” that are present in serum before viral infection. Natural antibodies have been defined as antibodies that are produced without any previous infection, vaccination, other foreign antigen exposure or passive immunization.
These antibodies can activate the classical complement pathway leading to lysis of enveloped virus particles long before the adaptive immune response is activated.
Many natural antibodies are directed against the disaccharide galactose α(1,3)-galactose (α-Gal), which is found as a terminal sugar on glycosylated cell surface proteins, and generated in response to production of this sugar by bacteria contained in the human gut.
Rejection of xenotransplantated organs is thought to be, in part, the result of natural antibodies circulating in the serum of the recipient binding to α-Gal antigens expressed on the donor tissue
Immunoglobulin diversity
Virtually all microbes can trigger an antibody response. Successful recognition and eradication of many different types of microbes requires diversity among antibodies; their amino acid composition varies allowing them to interact with many different antigens.
It has been estimated that humans generate about 10 billion different antibodies, each capable of binding a distinct epitope of an antigen.
Although a huge repertoire of different antibodies is generated in a single individual, the number of genes available to make these proteins is limited by the size of the human genome.
Several complex genetic mechanisms have evolved that allow vertebrate B cells to generate a diverse pool of antibodies from a relatively small number of antibody genes.
Domain variability
The chromosomal region that encodes an antibody is large and contains several distinct gene loci for each domain of the antibody—the chromosome region containing heavy chain genes (IGH@) is found on chromosome 14, and the loci containing lambda and kappa light chain genes (IGL@ and IGK@) are found on chromosomes 22 and 2 in humans.
One of these domains is called the variable domain, which is present in each heavy and light chain of every antibody, but can differ in different antibodies generated from distinct B cells.
Differences, between the variable domains, are located on three loops known as hypervariable regions (HV-1, HV-2 and HV-3) or complementarity-determining regions (CDR1, CDR2 and CDR3). CDRs are supported within the variable domains by conserved framework regions.
The heavy chain locus contains about 65 different variable domain genes that all differ in their CDRs. Combining these genes with an array of genes for other domains of the antibody generates a large cavalry of antibodies with a high degree of variability.
This combination is called V(D)J recombination discussed below.
V(D)J recombination
Somatic recombination of immunoglobulins, also known as V(D)J recombination, involves the generation of a unique immunoglobulin variable region.
The variable region of each immunoglobulin heavy or light chain is encoded in several pieces—known as gene segments (subgenes). These segments are called variable (V), diversity (D) and joining (J) segments.
V, D and J segments are found in Ig heavy chains, but only V and J segments are found in Ig light chains. Multiple copies of the V, D and J gene segments exist, and are tandemly arranged in the genomes of mammals. In the bone marrow, each developing B cell will assemble an immunoglobulin variable region by randomly selecting and combining one V, one D and one J gene segment (or one V and one J segment in the light chain).
As there are multiple copies of each type of gene segment, and different combinations of gene segments can be used to generate each immunoglobulin variable region, this process generates a huge number of antibodies, each with different paratopes, and thus different antigen specificities.
The rearrangement of several subgenes (i.e. V2 family) for lambda light chain immunoglobulin is coupled with the activation of microRNA miR-650, which further influences biology of B-cells.
RAG proteins play an important role with V(D)J recombination in cutting DNA at a particular region.Without the presence of these proteins, V(D)J recombination would not occur.
After a B cell produces a functional immunoglobulin gene during V(D)J recombination, it cannot express any other variable region (a process known as allelic exclusion) thus each B cell can produce antibodies containing only one kind of variable chain.
Somatic hypermutation and affinity maturation
Following activation with antigen, B cells begin to proliferate rapidly. In these rapidly dividing cells, the genes encoding the variable domains of the heavy and light chains undergo a high rate of point mutation, by a process called somatic hypermutation (SHM).
SHM results in approximately one nucleotide change per variable gene, per cell division.As a consequence, any daughter B cells will acquire slight amino acid differences in the variable domains of their antibody chains.
This serves to increase the diversity of the antibody pool and impacts the antibody’s antigen-binding affinity.
Some point mutations will result in the production of antibodies that have a weaker interaction (low affinity) with their antigen than the original antibody, and some mutations will generate antibodies with a stronger interaction (high affinity).
B cells that express high affinity antibodies on their surface will receive a strong survival signal during interactions with other cells, whereas those with low affinity antibodies will not, and will die by apoptosis.
Thus, B cells expressing antibodies with a higher affinity for the antigen will outcompete those with weaker affinities for function and survival allowing the average affinity of antibodies to increase over time.
The process of generating antibodies with increased binding affinities is called affinity maturation. Affinity maturation occurs in mature B cells after V(D)J recombination, and is dependent on help from helper T cells.
Class switching
Isotype or class switching is a biological process occurring after activation of the B cell, which allows the cell to produce different classes of antibody (IgA, IgE, or IgG).
The different classes of antibody, and thus effector functions, are defined by the constant (C) regions of the immunoglobulin heavy chain.
Initially, naive B cells express only cell-surface IgM and IgD with identical antigen binding regions. Each isotype is adapted for a distinct function; therefore, after activation, an antibody with an IgG, IgA, or IgE effector function might be required to effectively eliminate an antigen.
Class switching allows different daughter cells from the same activated B cell to produce antibodies of different isotypes.
Only the constant region of the antibody heavy chain changes during class switching; the variable regions, and therefore antigen specificity, remain unchanged.
Thus the progeny of a single B cell can produce antibodies, all specific for the same antigen, but with the ability to produce the effector function appropriate for each antigenic challenge.
Class switching is triggered by cytokines; the isotype generated depends on which cytokines are present in the B cell environment.
Class switching occurs in the heavy chain gene locus by a mechanism called class switch recombination (CSR). This mechanism relies on conserved nucleotide motifs, called switch (S) regions, found in DNA upstream of each constant region gene (except in the δ-chain).
The DNA strand is broken by the activity of a series of enzymes at two selected S-regions.
The variable domain exon is rejoined through a process called non-homologous end joining (NHEJ) to the desired constant region (γ, α or ε). This process results in an immunoglobulin gene that encodes an antibody of a different isotype.
Specificity designations
An antibody can be called monospecific if it has specificity for the same antigen or epitope,or bispecific if they have affinity for two different antigens or two different epitopes on the same antigen.
A group of antibodies can be called polyvalent (or unspecific) if they have affinity for various antigens or microorganisms.Intravenous immunoglobulin, if not otherwise noted, consists of a variety of different IgG (polyclonal IgG). In contrast, monoclonal antibodies are identical antibodies produced by a single B cell.
Asymmetrical antibodies
Heterodimeric antibodies, which are also asymmetrical and antibodies, allow for greater flexibility and new formats for attaching a variety of drugs to the antibody arms.
One of the general formats for a heterodimeric antibody is the “knobs-into-holes” format. This format is specific to the heavy chain part of the constant region in antibodies.
The “knobs” part is engineered by replacing a small amino acid with a larger one. It fits into the “hole”, which is engineered by replacing a large amino acid with a smaller one.
What connects the “knobs” to the “holes” are the disulfide bonds between each chain. The “knobs-into-holes” shape facilitates antibody dependent cell mediated cytotoxicity.
Single chain variable fragments (scFv) are connected to the variable domain of the heavy and light chain via a short linker peptide. The linker is rich in glycine, which gives it more flexibility, and serine/threonine, which gives it specificity.
Two different scFv fragments can be connected together, via a hinge region, to the constant domain of the heavy chain or the constant domain of the light chain.This gives the antibody bispecificity, allowing for the binding specificities of two different antigens.
The “knobs-into-holes” format enhances heterodimer formation but doesn’t suppress homodimer formation.
To further improve the function of heterodimeric antibodies, many scientists are looking towards artificial constructs.
Artificial antibodies are largely diverse protein motifs that use the functional strategy of the antibody molecule, but aren’t limited by the loop and framework structural constraints of the natural antibody.
Being able to control the combinational design of the sequence and three-dimensional space could transcend the natural design and allow for the attachment of different combinations of drugs to the arms.
Heterodimeric antibodies have a greater range in shapes they can take and the drugs that are attached to the arms don’t have to be the same on each arm, allowing for different combinations of drugs to be used in cancer treatment.
Pharmaceuticals are able to produce highly functional bispecific, and even multispecific, antibodies. The degree to which they can function is impressive given that such a change of shape from the natural form should lead to decreased functionality.
Medical applications
Disease diagnosis
Detection of particular antibodies is a very common form of medical diagnostics, and applications such as serology depend on these methods.
For example, in biochemical assays for disease diagnosis,a titer of antibodies directed against Epstein-Barr virus or Lyme disease is estimated from the blood.
If those antibodies are not present, either the person is not infected or the infection occurred a very long time ago, and the B cells generating these specific antibodies have naturally decayed.

medical diagnosis of antibodies
In clinical immunology, levels of individual classes of immunoglobulins are measured by nephelometry (or turbidimetry) to characterize the antibody profile of patient.Elevations in different classes of immunoglobulins are sometimes useful in determining the cause of liver damage in patients for whom the diagnosis is unclear.[1] For example, elevated IgA indicates alcoholic cirrhosis, elevated IgM indicates viral hepatitis and primary biliary cirrhosis, while IgG is elevated in viral hepatitis, autoimmune hepatitis and cirrhosis.
Autoimmune disorders can often be traced to antibodies that bind the body’s own epitopes; many can be detected through blood tests. Antibodies directed against red blood cell surface antigens in immune mediated hemolytic anemia are detected with the Coombs test.The Coombs test is also used for antibody screening in blood transfusion preparation and also for antibody screening in antenatal women.
Practically, several immunodiagnostic methods based on detection of complex antigen-antibody are used to diagnose infectious diseases, for example ELISA, immunofluorescence, Westernblot, immunodiffusion, immunoelectrophoresis, and magnetic immunoassay.
Antibodies raised against human chorionic gonadotropin are used in over the counter pregnancy tests.
New dioxaborolane chemistry enables radioactive fluoride (18F) labeling of antibodies, which allows for positron emission tomography (PET) imaging of cancer.
Disease therapy
Targeted monoclonal antibody therapy is employed to treat diseases such as rheumatoid arthritis,multiple sclerosis,psoriasis,and many forms of cancer including non-Hodgkin’s lymphoma,colorectal cancer, head and neck cancer and breast cancer.
Some immune deficiencies, such as X-linked agammaglobulinemia and hypogammaglobulinemia, result in partial or complete lack of antibodies.These diseases are often treated by inducing a short term form of immunity called passive immunity. Passive immunity is achieved through the transfer of ready-made antibodies in the form of human or animal serum, pooled immunoglobulin or monoclonal antibodies, into the affected individual.
Prenatal therapy
Rh factor, also known as Rh D antigen, is an antigen found on red blood cells; individuals that are Rh-positive (Rh+) have this antigen on their red blood cells and individuals that are Rh-negative (Rh–) do not.
During normal childbirth, delivery trauma or complications during pregnancy, blood from a fetus can enter the mother’s system.
In the case of an Rh-incompatible mother and child, consequential blood mixing may sensitize an Rh- mother to the Rh antigen on the blood cells of the Rh+ child, putting the remainder of the pregnancy, and any subsequent pregnancies, at risk for hemolytic disease of the newborn.
Rho(D) immune globulin antibodies are specific for human RhD antigen.Anti-RhD antibodies are administered as part of a prenatal treatment regimen to prevent sensitization that may occur when a Rh-negative mother has a Rh-positive fetus.
Treatment of a mother with Anti-RhD antibodies prior to and immediately after trauma and delivery destroys Rh antigen in the mother’s system from the fetus.
It is important to note that this occurs before the antigen can stimulate maternal B cells to “remember” Rh antigen by generating memory B cells.
Therefore, her humoral immune system will not make anti-Rh antibodies, and will not attack the Rh antigens of the current or subsequent babies.
Rho(D) Immune Globulin treatment prevents sensitization that can lead to Rh disease, but does not prevent or treat the underlying disease itself.
Credit:
https://en.wikipedia.org/wiki/Antibody#Forms
Leave an answer
You must login or register to add a new answer.